As Hominins Continued to Evolve Their Limbs
Introduction
Darwin's great insight, and the unifying principle of biology today, is that all species are related to one another like sisters, cousins, and distant kin in a vast family tree of life. The implications are breathtaking; if we could travel back far enough in time, we would find common ancestors between ourselves and every other living organism, from porcupines to flamingoes to cactuses. Our immediate evolutionary family is comprised of the hominoids, the group of primates that includes the "lesser apes" (siamangs and gibbons) as well as the "great apes" (chimpanzees, bonobos, gorillas, and orangutans). Among the great apes, our closest relatives are the chimpanzees and bonobos (Figure 1). The fossil record, along with studies of human and ape DNA, indicate that humans shared a common ancestor with chimpanzees and bonobos sometime around 6 million years ago (mya). We begin this discussion of our species' evolution in Africa, near the end of the geological time period known as the Miocene, just before our lineage diverged from that of chimpanzees and bonobos.
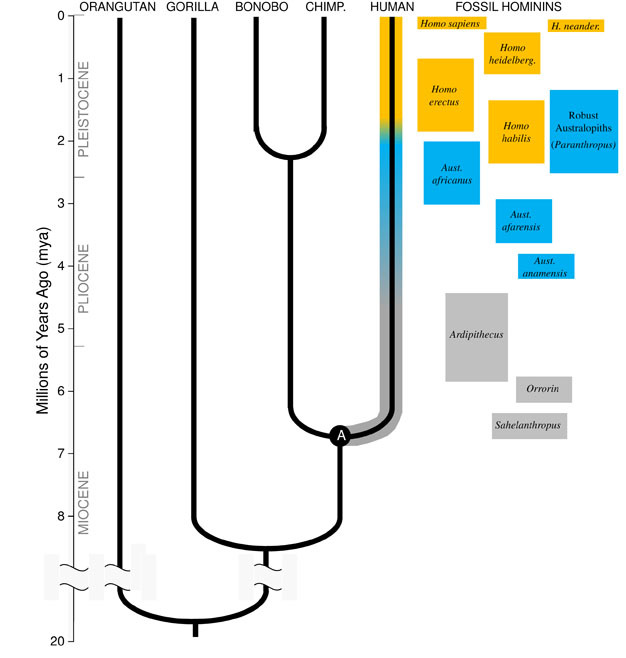
Figure 1: The human evolutionary family tree.
Relationships and estimated divergence times of the living apes are shown. Hominins are all species, including side-branches and extinct species, on the human line (highlighted) after our Last Common Ancestor with chimpanzees & bonobos (marked "A"). Some fossil hominin species are shown in the right-hand column, with their approximate age ranges indicated; early hominins: gray, Australopithecus: blue, Homo: orange.
Miocene Origins of the Hominin Lineage
In order to understand the evolution of any species, we must first establish its ancestral state: what sort of animal did it evolve from? For our lineage, this requires that we try and reconstruct the Last Common Ancestor of humans and chimpanzees (marked "A" in Figure 1). The Human-Chimpanzee Last Common Ancestor (HC-LCA) is the species from which the hominin lineage and the chimpanzee & bonobo lineage diverged. Hominins are species on our branch of the hominoid tree after the split with the chimpanzee & bonobo line, including all of the extinct species and evolutionary side branches (Figure 1).
There was a great diversity of ape species in the Miocene, with dozens of species known from the fossil record across Africa, Europe, and Asia. These species varied in their anatomy and ecology, and it is not clear which, if any, of the fossil species discovered thus far represent the HC-LCA (Kunimatsu et al. 2007; Young and MacLatchy, 2004). Nonetheless, we know from fossil and comparative evidence that it was much more similar to living apes than to living humans. The HC-LCA would have had an ape-sized brain and body, with relatively long arms and fingers and a grasping foot that allowed it to forage in the trees. The canine teeth were probably large and sharp, as seen in several Miocene hominoids. Moreover, the canines were probably sexually dimorphic, with males having much larger canines than females, as seen among the living great apes and Miocene fossils. Like living apes it would have walked quadrupedally (on all fours) when on the ground, and its diet would have consisted almost entirely of plant foods, primarily fruit and leaves.
Early Hominins
Changes from an ape-like anatomy are discernible in hominoid fossils from the late Miocene in Africa. Some hominoid species from this period exhibit traits that are typical of humans but are not seen in the other living apes, leading paleoanthropologists to infer that these fossils represent early members of the hominin lineage. The first human-like traits to appear in the hominin fossil record are bipedal walking and smaller, blunt canines.
The oldest hominins currently known are Sahelanthropus tchadensis from Chad (Brunet et al. 2005) and Orrorin tugenensis from Kenya (Senut et al. 2001). Sahelanthropus, dated to between 6 and 7 mya, is known from a largely complete skull and some other fragmentary remains. Its brain size, 360cc, is within the range seen in chimpanzees, and the skull has a massive brow ridge, similar in thickness to male gorillas (Brunet et al. 2005). However, the position and orientation of the foramen magnum, the hole in the base of the skull through which the spinal cord passes, suggests that Sahelanthropus stood and walked bipedally, with its spinal column held vertically as in modern humans rather than horizontally as in apes and other quadrupeds (Zollikofer et al. 2005). Orrorin is known primarily from postcranial fossils, including a partial femur. The proximal portion of the femur shows similarities to those of modern humans, suggesting the species was bipedal (Pickford et al. 2002). No skulls of Orrorin have been recovered, and so its cranial morphology and brain size are uncertain. In both Orrorin and Sahelanthropus the canine teeth of males are larger and more pointed than in modern humans, but are small and blunt compared to the canines of male apes. This suggests that canine sexual dimorphism — and by extension, competition among males for mating access to females — was diminished in these early hominins compared to the great apes.
By far the best known early hominin is Ardipithecus ramidus, a 4.4 million year old species from Ethiopia, which is known from a nearly complete skeleton as well as numerous other dental and skeletal remains (White et al. 2009). Ar. ramidus and an older, related species known from fragmentary remains, Ar. kadabba (5.8–5.2 mya), have reduced canines similar to those of Orrorin and Sahelanthropus. The skull of Ar. ramidus is rather ape-like and broadly similar to that of Sahelanthropus, with a small chimpanzee-sized brain of 300–350cc (Figure 2). The Ardipithecus postcranial skeleton is intriguing. Although badly fragmented, the pelvis recovered reveals a morphology quite different from that of living apes, with a shorter, more bowl-like shape that strongly suggests Ardipithecus walked bipedally; this is consistent with the foramen magnum position, which suggests an upright posture. However, its long forelimbs and fingers and its divergent, grasping first toe (hallux) suggest Ardipithecus spent much of its time in the trees. The overall impression is of a largely arboreal species that walked bipedally whenever it ventured to the ground.
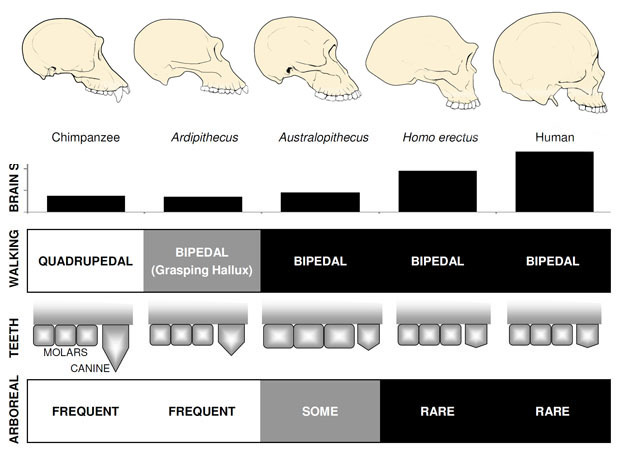
Figure 2: Anatomical comparisons of apes, early hominins, Australopithecus, Homo erectus, and humans.
A male chimpanzee skull is shown as an example of modern apes. Early hominins and Australopithecus retained ape-size brains. Bipedal walking evolved very early in the hominin lineage, but Ardipithecus (and possibly other early hominins) retained a grasping foot that may have diminished bipedal efficiency. Reduced canine size also evolved early in the hominin lineage, although early hominin canines were larger and more pointed than those of later hominins. Molar size increased with Australopithecus but later is reduced in Homo (note: premolars are not shown in this schematic). Arboreal locomotion, as indicated by the presence of long arms, curved fingers and toes, and other forelimb features, was common throughout much of the hominin lineage.
Australopithecus
Around 4mya we find the earliest members of the genus Australopithecus, hominins which were adept terrestrial bipeds but continued to use the trees for food and protection. The first specimens of Australopithecus were discovered in South Africa in 1924 (Dart, 1925), and research efforts over the subsequent eight decades have produced hundreds of fossils from several species at sites all across East and Southern Africa. We now know that Australopithecus was a highly successful genus that persisted for nearly three million years (Figure 1).
The best-known Australopithecus species are A. afarensis (3.6–2.9 mya) from East Africa and A. africanus (3.2–2.0mya) from South Africa. The pelvis and lower limb of these species clearly indicates that they were fully bipedal: the pelvis is short and bowl-shaped, bringing the gluteal muscles around to the side of the body, as in modern humans, for trunk stabilization during bipedalism, and the first toe is in line with the other toes (Ward, 2002; Harcourt-Smith and Aiello, 2004). The Australopithecus foot may even have had a human-like arch, based on analysis of the metatarsals and the fossilized Laetoli footprints (Ward et al. 2011). Nonetheless, compared to modern humans, the forearms were long and the fingers and toes were long and somewhat curved, suggesting that Australopithecus regularly used the trees to forage and perhaps as a refuge from predators at night. This mixed terrestrial & arboreal strategy would have served these species well in the mixed woodland and savannah environments they inhabited.
Brain size in Australopithecus ranged between 390 and 515cc, similar to chimpanzees and gorillas (Falk et al. 2000), suggesting cognitive abilities were broadly similar to living apes (Figure 2). Body size in Australopithecus was rather small and sexually dimorphic, about 30kg for females and 40kg for males (McHenry, 1992). This level of dimorphism is not reflected in the canines, which were small, blunt, and monomorphic as in earlier hominins.
Unlike the canines, molar teeth in Australopithecus were much larger than those of earlier hominins, and had thicker enamel. This suggests their diet included hard, low quality plant foods that required powerful chewing to process. A subgroup of Australopithecus, known as the "robust" australopiths (often labeled by a separate genus Paranthropus) because of their enormous teeth and chewing muscles, took this adaptation to the extreme. Most Australopithecus species were extinct by 2 mya, but some robust forms persisted until about 1.2 mya in East and South Africa.
The Genus Homo
The earliest fossils of our own genus, Homo, are found in East Africa and dated to 2.3 mya (Kimbel et al. 1997). These early specimens are similar in brain and body size to Australopithecus, but show differences in their molar teeth, suggesting a change in diet. Indeed, by at least 1.8 mya, early members of our genus were using primitive stone tools to butcher animal carcasses, adding energy-rich meat and bone marrow to their plant-based diet.
The oldest member of the genus Homo, H. habilis (2.3–1.4 mya) is found in East Africa and is associated with butchered animal bones and simple stone tools (Blumenschine et al. 2003). Its more formidable and widespread descendant, H. erectus, is found throughout Africa and Eurasia and persisted from 1.9 mya to 100 kya, and perhaps even later (Anton, 2003). Like modern humans, H. erectus lacked the forelimb adaptations for climbing seen in Australopithecus (Figure 2). Its global expansion suggests H. erectus was ecologically flexible, with the cognitive capacity to adapt and thrive in vastly different environments. Not surprisingly, it is with H. erectus that we begin to see a major increase in brain size, up to 1,250cc for later Asian specimens (Anton, 2003). Molar size is reduced in H. erectus relative to Australopithecus, reflecting its softer, richer diet.
Around 700 kya, and perhaps earlier, H. erectus in Africa gave rise to H. heidelbergensis, a species very much like us in terms of body proportions, dental adaptations, and cognitive ability (Rightmire, 2009). H. heidelbergensis, often referred to as an "archaic" Homo sapiens, was an active big-game hunter, produced sophisticated Levallois style tools, and by at least 400 kya had learned to control fire (Roebroeks and Villa, 2011). Neanderthals (H. neanderthalensis), cold-adapted hominins with stout physiques, complex behaviors, and brains similar in size to ours, are thought to have evolved from H. heidelbergensis populations in Europe by at least 250 kya (Rightmire, 2008; Hublin, 2009).
Fossil and DNA evidence suggest our own species, H. sapiens, evolved in Africa 200 kya (Relethford, 2008; Rightmire, 2009), probably from H. heidelbergensis. The increased behavioral sophistication of H. sapiens, as indicated by our large brains (1,400cc) and archeological evidence of a broader tool set and clever hunting techniques, allowed our species to flourish and grow on the African continent. By 100kya, our species spilled into Eurasia, eventually expanding across the entire globe into Australia and the Americas (DiGiorgio et al. 2009). Along the way our species displaced other hominins they encountered, including Neanderthals in Europe and similar forms in Asia. (Note that not all agree with this interpretation of the data, see Tryon and Bailey). Studies of ancient DNA extracted from Neanderthal fossils suggest our species may have occasionally interbred with them (Green et al., 2010). Our increasing global impact continues today, as cultural innovations such as agriculture and urbanization shape the landscape and species around us.
Summary
The evolution of our species from an ape-like Miocene ancestor was a complex process. Our lineage is full of side branches and evolutionary dead ends, with species like the robust australopiths that persisted for over a million years before fading away. Some human traits, like bipedalism, evolved very early, while others, like large brains, did not evolve until relatively recently. Still other traits, like molar size, evolved in one direction only to be pushed back later by changing ecological pressures. Rather than a powerful ship charting a straight course toward some pre-determined destination, the evolution of our lineage — indeed, of any species' lineage — fits the image of a lifeboat tossed about by the shifting seas of environmental change, genetic luck, and geological chance. One wonders where the next six million years might take us.
Source: https://www.nature.com/scitable/knowledge/library/overview-of-hominin-evolution-89010983/
{kind=link}
Post a Comment for "As Hominins Continued to Evolve Their Limbs"